


Initialising ...
Initialising ...
Initialising ...
Initialising ...
Initialising ...
Initialising ...
Initialising ...
樋口 真理子; Pinak, M.
no journal, ,
放射線により損傷が10から20ベースペア以内に複数生じることがあり、これをクラスター損傷と呼ぶ。修復酵素hOGG1はDNA上の損傷残基である8オキソグアニンを修復する酵素だが、8オキソグアニンの相補鎖上の数ベースペア離れた位置に一本鎖鎖切断がある場合、修復が阻害されることが実験によりわかっている。8オキソグアニンと他の損傷の相対位置により修復率が変わるが、その原因は明確ではない。本研究では、クラスター損傷DNAと修復酵素hOGG1の分子動力学シミュレーションを行い、結合状態について調べた。クラスター損傷のモデルは8オキソグアニンと一本鎖切断を含み、二つの損傷の相対位置を変えて、それぞれシミュレーションを行った。その結果、8オキソグアニンからみて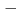 3ベースペアに鎖切断があると、+3ベースペアにある場合と比較して結合力が低くなる結果が得られた。この結果は、クラスター損傷を持つDNAと酵素hOGG1との結合力が、損傷の修復率に反映していることを支持する。
3ベースペアに鎖切断があると、+3ベースペアにある場合と比較して結合力が低くなる結果が得られた。この結果は、クラスター損傷を持つDNAと酵素hOGG1との結合力が、損傷の修復率に反映していることを支持する。
松本 富美子; 安達 基泰; 清水 瑠美; 目黒 瑞枝; 玉田 太郎; 加藤 尚志; 黒木 良太
no journal, ,
トロンボポエチン(TPO)は、血液細胞の一つである巨核球を増殖させるとともに血小板への分化を刺激するサイトカインである。そのためTPOによる受容体活性化機構に関する知見は学術的に重要であるだけでなく、創薬研究にも重要な情報を与える。われわれはecTPORのシステイン残基を選択的に変異させることにより、非特異的なジスルフィド結合形成の抑制を試みた。さらに、ecTPORのTPO結合部位の同定を行うためにecTPOR中に存在する2つのCRH領域(CRH-1とCRH-2)それぞれの発現系を構築した。大腸菌により発現したCRH-1はニッケルカラムにより粗精製した後リフォールディングを行い、ゲル濾過カラムにより精製することができた。CRH-1とTPO活性領域との相互作用は、ゲル濾過と光散乱を用いて解析した。その結果、CRH-1はTPOと親和性を有することを見いだした。先に動物細胞で発現させたecTPORにはTPO結合部位が一か所だけ存在することを明らかにしている。今回の結果からecTPOR中のTPO相互作用部位はCRH-1であると結論できる。
玉田 太郎; 木下 誉富*; 山田 貢; 栗原 和男; 大原 高志; 安達 基泰; 多田 俊治*; 黒木 良太
no journal, ,
エラスターゼは、蛋白質の立体構造に基づく薬物設計(SBDD)研究の代表的な題材として用いられてきたセリンプロテアーゼである。エラスターゼの全原子構造情報取得によってセリンプロテアーゼの触媒機構解明を目指すとともに、SBDD研究に供するため、ブタ膵臓由来のエラスターゼ(PPE)の単独及び阻害剤複合体での中性子結晶構造解析を実施した。中性子回折実験はJRR3に設置された生体高分子用回折計BIX-3において実施し、同一結晶を用いた取得したX線回折データを相補的に用いた構造精密化により高精度の全原子構造情報を取得した。PPE単独(1.9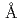 分解能)、及びエラスターゼの基質を模倣したペプチド様の阻害剤との複合体状態(1.65分解能)での中性子構造解析に成功した。今回取得した阻害剤結合前・後の水素原子を含む全原子構造情報は、これまでにセリンプロテアーゼの触媒機構において議論されてきた反応開始時の活性化制御機構や遷移状態における低障壁水素結合の存在や酸素陰イオンホールの形成について重要な知見を与えると同時に、活性部位の水和構造の変化についても重要な知見を与えた。
分解能)、及びエラスターゼの基質を模倣したペプチド様の阻害剤との複合体状態(1.65分解能)での中性子構造解析に成功した。今回取得した阻害剤結合前・後の水素原子を含む全原子構造情報は、これまでにセリンプロテアーゼの触媒機構において議論されてきた反応開始時の活性化制御機構や遷移状態における低障壁水素結合の存在や酸素陰イオンホールの形成について重要な知見を与えると同時に、活性部位の水和構造の変化についても重要な知見を与えた。
安達 基泰; 玉田 太郎; 黒木 良太; 日高 興士*; 木村 徹*; 木曽 良明*; 山本 航平*; 城所 俊一*
no journal, ,
HIV-1プロテアーゼ(HIVPR)は、エイズ治療のための創薬標的タンパク質である。HIVPRに対する阻害剤設計においては、立体構造の特徴と阻害剤結合の速度論的・熱力学的解析から得られるパラメーターの相関を明らかにすることが重要である。本研究では、阻害剤結合による熱安定性の変化を指標として阻害剤結合を評価するため、プロトマー間でN及びC末端が隣接することに着目し、タンデムに結合した遺伝子間に2アミノ酸残基分のリンカー配列を挿入することで、一本鎖化HIVPRの作製を試みた。一本鎖化HIVPRは、野生型と同様に大腸菌内に封入体として発現した。リフォールディングした一本鎖化HIVPRを精製し、KNI-272との複合体を結晶化した。得られた結晶を用いて、放射光施設(SPring-8)にて1.3分解能の回折データを収集し、R値19%まで構造を精密化した。一本鎖化HIVPRの立体構造は、野生型とほとんど同じ(RMSD[C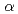 ]=0.17)であったが、挿入したリンカー部分の電子密度は観測されなかった。同試料を用いた阻害剤結合の評価結果も報告する。
]=0.17)であったが、挿入したリンカー部分の電子密度は観測されなかった。同試料を用いた阻害剤結合の評価結果も報告する。
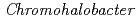 sp.560株由来
sp.560株由来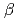 -LactamaseのX線結晶解析
-LactamaseのX線結晶解析新井 栄揮; 米澤 悌; 岡崎 伸生; 玉田 太郎; 徳永 廣子*; 石橋 松二郎*; 徳永 正雄*; 黒木 良太
no journal, ,
近年、希少金属の供給の不安定化に伴って、海水等から希少金属イオンを効率的に回収する技術の確立が求められている。われわれは、分子表面に数多くの負電荷を有する好塩性蛋白質を、希少金属を捕集する材料として着目し、数種類の好塩性蛋白質のX線結晶解析を実施し、無機イオンの結合にかかわる蛋白質分子表面の構造学的特徴(極性原子の配置や配位数など)の解析を行っている。その研究の一環として、われわれは中度好塩菌 sp.560由来・好塩性-LactamaseのX線結晶解析を行い、立体構造決定に成功した。現在、構造解析結果について極性原子の配置や配位数などを精査し、Na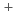 やMg
やMg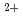 などが結合しうる部位の抽出を行っている。これらの金属結合部位に変異導入による電荷や構造的な摂動を与えて無機イオン選択性を操作できれば、希少金属結合部位を人工的に創製することも可能になると考えられる。この技術が確立されれば、原子力発電所の事故により漏出した放射性セシウム等の回収にも応用できると期待される。
などが結合しうる部位の抽出を行っている。これらの金属結合部位に変異導入による電荷や構造的な摂動を与えて無機イオン選択性を操作できれば、希少金属結合部位を人工的に創製することも可能になると考えられる。この技術が確立されれば、原子力発電所の事故により漏出した放射性セシウム等の回収にも応用できると期待される。
有森 貴夫; 岡崎 伸生; 中澤 昌美*; 宮武 和孝*; 上田 光宏*; 玉田 太郎
no journal, ,
好熱性細菌由来キチン分解酵素Ra-ChiCは、キチン結合ドメインと触媒ドメインから構成されており、その触媒ドメインはガチョウ型リゾチームと18%程度のアミノ酸相同性を有している。しかしながら、Ra-ChiCはキチナーゼ活性のみを示し、リゾチーム活性は示さない。そこでわれわれはRa-ChiCがいかにして有為なキチナーゼ活性を発揮するかを理解することを目的とし、Ra-ChiCの立体構造解析を行った。その結果、ガチョウ型リゾチームの触媒残基であるGlu残基は、Ra-ChiCにおいて側鎖を含め立体的によく保存されていた(Glu141)が、ガチョウ型リゾチームで活性に関与していると考えられている2つのAsp残基については、Ra-ChiCでは1つは立体的にほぼ似た箇所にGluとして存在し(Glu162)、もう1つは一次構造上全く異なる位置にあるAsp残基が三次構造上で近い位置に存在していた(Asp226)。さらに、Ra-ChiCではAsp226が存在するループ領域が活性部位を覆うような構造をしており、ガチョウ型リゾチームと比較して基質結合ポケットを狭めていた。これらの違いがキチナーゼ活性とリゾチーム活性の違いを創出していると考えられた。
清水 瑠美; 松本 富美子; 新井 栄揮; 大原 高志; 安達 基泰; 玉田 太郎; 黒木 良太; 西宮 佳志*; 近藤 英昌*; 津田 栄*
no journal, ,
不凍タンパク質(AFP)は、氷の表面に結合することで氷結晶の成長を抑制し、体液の凝固点を下げる働きを持つタンパク質である。AFPは、非常にユニークな機能を持つことから、食品,医療などさまざまな分野での産業利用が期待されている。北海道沿岸に生息するナガガジの体内では多数のアイソフォームが発現しており、これらはSP型及びQAE型に分類される。われわれは、活性が低いSP型のnfeAFP6と活性が高いQAE型のnfeAFP8の間で、アミノ酸配列が異なる部分に着目し、キメラ体4種類と部位特異的変異体14種類を大腸菌発現系により調製して、氷結晶成長抑制活性を比較した。その結果、nfeAFP6の分子表面に存在するAla19一か所のみをValに置換したnfeAFP6 A19V変異体がQAE型と同様の高い氷結晶成長抑制作用を持つことを見いだした。さらに氷結晶の成長抑制活性と分子構造の安定性の相関を検討するために、各変異体の構造安定性を円偏光二色性の温度依存性から比較した。その結果、nfeAFP6 A19V変異体の構造安定性は、野生型よりも低く、その他の変異体に関して活性の強さと構造安定性に相関は見られなかった。よって、変異型AFPの不凍活性変化は、その安定性の変化に由来するのではなく、変異部位の導入によって、水和構造を変化させたからであると考えられる。
-ラクタマーゼToho-1の1.5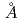 分解能データ取得に成功
分解能データ取得に成功栗原 和男; 玉田 太郎; 大原 高志; 黒木 良太
no journal, ,
われわれは、生体高分子用中性子単結晶回折装置BIX-3, BIX-4(原子力機構・JRR-3)を用い、タンパク質等の生体高分子における水素・水和構造と機能の解明を行っている。現在、BIX-3, BIX-4の装置高度化を進めており、その一環として、両回折装置の試料環境に対し、独自設計の新規試料ゴニオメータと組合せた窒素吹付け型低温装置(到達温度:100K)を導入した。2010年度には、タンパク質大型結晶(2-3mm角)に対して本低温装置を適用した測定を本格的に開始した。下記のタンパク質がその最初の例である。-ラクタマーゼToho-1(共同研究:理研・宮野主任研究員ら)は、抗生物質の-ラクタム環を加水分解し不活性にする酵素の一種であり、バクテリアが抗生物質への耐性を示す原因物質の一つである。BIX-4を用い、そのR274N/R276N変異体結晶から、従来の常温での実験結果に基づいた格子定数と結晶体積から予想される分解能を上回る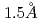 分解能の回折イメージ取得に成功した。これはこれまでのタンパク質中性子構造の中でも最高分解能に並ぶ。さらに、同一結晶を用いてX線回折データも収集した(SPring-8・BL38B1)。現在、中性子とX線の両回折データを用いた同時構造精密化を実施中である。今後、両試料の解析結果をもとに、-ラクタマーゼの認識・反応機構を解明し、阻害剤開発に有用な情報を得ることが期待できる。
分解能の回折イメージ取得に成功した。これはこれまでのタンパク質中性子構造の中でも最高分解能に並ぶ。さらに、同一結晶を用いてX線回折データも収集した(SPring-8・BL38B1)。現在、中性子とX線の両回折データを用いた同時構造精密化を実施中である。今後、両試料の解析結果をもとに、-ラクタマーゼの認識・反応機構を解明し、阻害剤開発に有用な情報を得ることが期待できる。
目黒 瑞枝; 安達 基泰; 黒木 良太; 谷崎 祐太*; 田原 彩香*; 別府 実穂*; 永澤 和道*; 加藤 尚志
no journal, ,
大腸菌組換えタンパク質の発現では、mRNAの安定性に由来する翻訳効率だけでなく、フォールディングの容易さや溶解性さらにはタンパク質分解酵素への抵抗性などタンパク質固有の性質にも着目する必要がある。われわれは、比較生物学的な観点から4本ヘリックスバンドル型サイトカインの構造と機能に着目している。これまでヒトのサイトカインのほか、両生類ツメガエル( )由来のエリスロポエチン(
)由来のエリスロポエチン(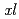 EPO),トロンボポエチン(TPO)の同定に成功し、ヒトEPO(hEPO)及びヒトトロンボポエチン(hTPO)との構造・機能の比較を行っている。EPOは、hEPOとの一次構造相同性が38%であり、またTPO, hTPOの相同性は28%である。しかし、これらのサイトカインを大腸菌で発現させた場合、EPOの発現量だけ著しく低かった。この原因としてEPOが大腸菌内で消化されている可能性に着目し、EPOのC末端のアミノ酸を置換又は数残基のアミノ酸を欠失・付加した変異体を作製し、各々の発現量を比較したところ、発現量が増大した変異体があった。特に、EPOのC末端に6残基のヒスチジンを付加した変異体が最も高い発現量を示した。この変異体を、これまでの精製系に適用したところ、最終精製物の収量が約8倍増加した。EPO発現量が向上したのは、C末端側への変異導入により、大腸菌タンパク質分解酵素からの分解を免れたためであると考えられる。
EPO),トロンボポエチン(TPO)の同定に成功し、ヒトEPO(hEPO)及びヒトトロンボポエチン(hTPO)との構造・機能の比較を行っている。EPOは、hEPOとの一次構造相同性が38%であり、またTPO, hTPOの相同性は28%である。しかし、これらのサイトカインを大腸菌で発現させた場合、EPOの発現量だけ著しく低かった。この原因としてEPOが大腸菌内で消化されている可能性に着目し、EPOのC末端のアミノ酸を置換又は数残基のアミノ酸を欠失・付加した変異体を作製し、各々の発現量を比較したところ、発現量が増大した変異体があった。特に、EPOのC末端に6残基のヒスチジンを付加した変異体が最も高い発現量を示した。この変異体を、これまでの精製系に適用したところ、最終精製物の収量が約8倍増加した。EPO発現量が向上したのは、C末端側への変異導入により、大腸菌タンパク質分解酵素からの分解を免れたためであると考えられる。